www.fimpolgy.com/JTF
The Journal of Theoretical Fimpology. Volume 1, Issue 2: e-20130512-1-2-4. May 18, 2013 (www.fimpology.com)
A Noninherited-Paternal-Antigen-Centric Model for the Birth Order and Sibship Size Effect in Transplantation Immunology
Shu-dong Yin
ORCID 0009-0005-8661-6889
Cory H. E. R. & C. Inc., Burnaby, British Columbia, Canada
The survival of grafts at the individual macroorganism level involves in a complex
tolerogenic mechanism. In humans, the relationship between the fetus, postnatal offspring,
mother and biological father has been revealed to be associated with tolerogenic
mechanisms in clinical transplantation, among which, the model of maternal-to-fetal cell
transmission mediated tolerance to noninherited maternal antigens (NIMAs) was once
highlighted. However, the NIMAs-centric theory along cannot explain the birth order
effect and the NIMA paradox. Recently, six models of pregnancy-associated eukaryotic
cell transmissions were summarized: fetus-to-maternal cell transmission (FMCT),
maternal-to-fetal cell transmission (MFCT), gestation-associated fetus-to-fetus cell
transmission (FFCT), breastmilk and breastfeeding-mediated maternal-to-infant cell
transmission (MICT), breastmilk and breastfeeding-mediated fetus-to-infant allogenic or
inter-sibling cell transmission (FICT), the fetus-breastmilk-breastfeeding-infant-cells
cycle (FBBIC Cycle). Based on these advances, I propose a novel theoretical model
called noninherited-paternal-antigen-centric model (NIPA-CM) for understanding the
birth order and sibship size effect in transplantation immunology. NIPA-CM illustrates
that FFCT and FICT may play a role in transferring noninherited paternal antigens
(NIPAs) to introduce sibling tolerance to them, and moreover, the variation of intersibling tolerance to NIPAs may depend on both birth order and sibship size.
Key Words: noninherited maternal antigens (NIMAs); noninherited paternal HLA antigens (NIPAs); birth order; sibship size; cell migration
Six decades ago, the concept of "acquired immunologic tolerance" was first proposed
by Billingham, Brent and Medawar [1-3]. In a retrospective study, Burlingham and
colleagues showed that the survival rates of kidneys from siblings expressing maternal
HLA antigens not inherited by the recipient were better than those of kidneys from
siblings expressing paternal HLA antigens not inherited by the recipient [4]. Moreover,
later studies on clinical cellular transplantation showed that blood and marrow stem cell
transplants from mother to child had better survival than that from paternal grafts [5,6].
Despite many unknowns and debates, this survival difference was partly accounted for by
the prenatal noninherited maternal antigens (NIMA)-induced tolerance theory [3,4,7-16].
However, the NIMA-centric theory cannot account for the birth order effect and the
NIMA paradox [4,14,17] in clinical transplantation.
During the past two decades, accumulating evidences from studies on humans and
non-human mammals have indicated that there are different eukaryotic cell transmissions
among mothers, fetuses and infants during gestation and lactation. Recently, six models
of eukaryotic cell transmission among mothers, fetuses and infants during gestation and
lactation have been summarized: (i) FMCT, (ii) MFCT, (iii) FFCT, (iv) MICT, (v) FICT,
and (vi) FBBIC Cycle [29]. Based on the this progress, I further propose a novel
theoretical model called noninherited-paternal-antigen (NIPA)-centric model (NIPA-CM)
for understanding the birth order and sibship size effect in transplantation immunology.
Hypothesis and Discussion
I. NIMA-centric model
The NIMA-induced tolerance theory was illustrated by Burlingham and colleagues in
their NIMA-centric model (a hypothesized family with two parents and three offspring)
[4], in which, it was assumed that the mother had alleles C and D of the gene for an HLA
antigen; and the father had alleles E and F of the same gene. According to Mendel's law
of inheritance, each offspring of them got one allele (C or D) from the mother and the
other (E or F) from the father, and therefore, the alleles for encoding the HLA antigen in
each offspring were one of the following four possibilities: CE, CF, DE and DF. If an
offspring got allele D from his or her mother, then C was called a noninherited maternal
allele, and the antigen encoded by C was described as a noninherited maternal antigen.
Similarly, if the offspring obtained E from his or her father, then F was called the
noninherited paternal allele, and the antigen encoded by F was noninherited paternal
antigen [4] (see Figure 1).
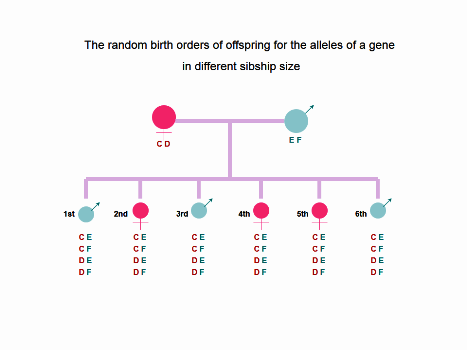
Figure 1. In a one-child family, the offspring has the four possibilities for the alleles of a gene: (1) CE, (2)
CF, (3) DE, and (4) DF. In a two-offspring family, the random birth orders of siblings for the alleles of a gene
has total following 16 possibilities: (1) CE+CE, (2) CE+CF, (3) CE+DE, (4) CE+DF, (5) CF+CE, (6) CF+CF, (7)
CF+DE, (8) CF+DF, (9) DE+CE, (10) DE+CF, (11) DE+DE, (12) DE+DF, (13) DF+CE, (14) DF+CF, (15) DF+DE, and
(16) DF+DF. As the sibship size increasing, the random birth orders of siblings for the alleles of a gene will
increase as the following: 64 possibilities in a three-offspring family, 256 possibilities in a four-offspring
family, 1024 possibilities in a five-offspring family, 4096 possibilities in a six-offspring family, and 4[N]
possibilities in a N-offspring family
In Burlingham's model, each offspring during his or her fetal life was hypothesized
not to have been exposed to the noninherited paternal HLA antigens (NIPAs) encoded by
the noninherited paternal alleles [4], which restricted its accounting for the clinical
phenomenon described by Opelz in 1990 that the survival of renal allografts donated by
mothers and fathers had no clinical difference [4], or later called "the NIMA Paradox" by
Burlingham and Benichou [17], and the birth order effect on the variation of inter-sibling
tolerance found by Bucher and colleagues in 2007 [14].
II. The birth order and sibship size effect
The sibship size and the birth order in epidemiological studies have been showed to
associate with allergic disorders, in which, asthma and atopy were found to be less
common in younger siblings than in elder siblings [18-22].
In recent years, siblings have increasingly gained attention as an important donors for
cellular and organ transplantation [12,23-26]. Bucher and colleagues first described the
phenomenon of the birth order effect on the variation of inter-sibling immune-tolerance
in a retrospective study on allogenic hematopoietic stem cell transplantation (HSCT), in
which, cellular grafts from younger siblings significantly decreased the risk of graftversus-host disease (GVHD) in recipients who were born as the first children of families
[14,27]. However, Gratwohl and colleagues later found no birth order effect in their
analysis of HLA-identical living sibling donor kidney transplantations [28].
III. The association of prenatal and postnatal eukaryotic cell transmission with NIMAs- tolerogenic and NIPAs- tolerogenic mechanisms
In normal pregnancies, fetus can come into contact with NIMAs via maternal fetal
molecular chimerism and maternal fetal cellular chimerism (the latter is also called the
maternal cell transmission to the fetus, MFCT), which is believed to produce immune
tolerance to NIMAs in the offspring. In contrast, the father does not have such advantages
in allowing the fetus to be exposed to NIPAs to produce immune tolerance to NIPAs
except delivering DNA molecular information to the offspring via Mendel’s Hereditary
Law.
So far, two tolerogenic mechanisms at the cellular level have been discussed: one is
the deletion of alloantigen-reactive T cells [13,30,31]; and the other is the development of
tolerogenic fetal T regulatory cells (Tregs) during gestation [3,31,32]. A few studies have
revealed that there were no differences in the frequencies of cytotoxic T cell precursors
(CTLp) and IL-2-producing helper T cell precursors (HTLp) against the noninherited
maternal and the paternal HLA antigens [10], or cell-mediated lympholysis response [9].
In regards to exposure to NIMAs, the offspring can come into contact with NIMAs via
both prenatal and postnatal mechanisms. For example, in HLA-A2 negative babies born
to HLA-A2+ mothers, soluble HLA-A2 (sHLA-A2) is a type of noninherited maternal
antigen (NIMAs), and such NIMAs can be exposed not only by HLA-A2 negative fetus
during pregnancy, which was confirmed by the detection of NIMA HLA-A2 in cord
blood, but also exposed postnatally via breastfeeding through the detection of NIMA
HLA-A2 in human milk from HLA-A2+ mothers [37]. It is clear that exposure to
noninherited maternal HLA antigens may also occur during lactation via both molecule
exchange at the molecular level and maternal-infant microchimerism at the cellular level,
while the infant is breastfeeding with milk from his or her own biological mother [37,38].
Recently, van Rood and colleagues proposed another possible mechanism involving
the microchimerism of maternal memory T cells anti-inherited paternal antigens (IPAs)
of the fetus [27]. In fact, the possibility of exposure to NIPAs during gestation via
prenatal FFCT has been suggested in a few of studies [14,33,34-36]. Moreover, paternal
alloantigens-tolerogenic fetal and maternal T regulatory cells (PATFM Tregs) were
hypothesized to play a role in postnatal antipaternal immune reaction to paternal antigens
[29].
Theoretically, fetus and/or infant actually can also obtain tolerance to NIPAs through
the following four mechanisms: (1) residual fetal cells from previous pregnancies in the
The Journal of Theoretical Fimpology. Volume 1, Issue 2: e-20130512-1-2-4. May 18, 2013 www.fimpology.com
Copyright © 2013-2023 by Cory H. E. R. & C. Inc. All Rights Reserved.
5
maternal body can migrate into the fetal body of next pregnancy via FFCT, regardless of
whether previous pregnancies were premature or matured, and whether they were aborted
or completed normally; (2) residual fetal cells from previous pregnancies enter the body
of the suckling infant via breastmilk and breastfeeding-mediated FICT; (3) FBBIC Cycle,
in which fetal cells of the current pregnancy may be prenatally exposed to NIPAs and
regained postnatally by the infant him- or her-self via breastfeeding; and (4) tolerogenic
maternal regulatory T cells to NIPA via prenatal MFCT and postnatal MICT.
IV. The birth order and sibship size-related NIPA-centric model
Based on Burlingham's NIMA-centric model [4], I propose a model called "the
noninherited paternal antigens-centric model" (NIPA-CM), in which, the mother in the
hypothesized family is assumed to have had no silent or identified abortions before
delivering her first living offspring. Both NIMAs and NIPAs are hypothesized to be
exposed to offspring during gestation and lactation via MFCT, FFCT, MICT, FICT, and
FBBIC Cycle.
Surprisingly, the coexistence of the birth order effect and the sibship size effect on the
variation of inter-sibling tolerance emerges, which may theoretically account for "the
NIMA Paradox". Interestingly, in this birth order and sibship size-related NIPA model,
the exposure to NIMAs via prenatal MFCT and FFCT and postnatal MICT, FICT and
FBBIC Cycle dose not show any birth order and/or sibship size effect because each
offspring has opportunities to contact NIMAs via both prenatal and postnatal multiple
routes (see Figure 2).
Generally speaking, in single pregnancy mammals, prenatal FFCT and MFCT and
postnatal FICT, MICT and FBBIC Cycle may offer later-born siblings the chance to be
exposed to NIPAs carried by their elder siblings or previously aborted embryos/fetuses,
and therefore, to induce tolerance to NIPA. In contrast, the elder siblings have no chance
to contact NIPAs carried by their younger siblings and therefore no yielded tolerance to
NIPAs carried by younger siblings.
Theoretically, the survival rate of grafts from older siblings in younger sibling
recipients is usually higher than that of grafts from younger siblings in elder sibling
recipients, meaning less graft rejection in younger sibling recipients, and less graftversus-host disease (GVHD) in elder sibling recipients.
In the NIPAs-centric model (see Figure 2), each younger siblings may have both
prenatal and postnatal opportunities to expose to NIPAs carried by the first child in a
family, but the eldest sibling has no chance to expose to NIPAs carried by his or her
younger siblings, which may account for Bucher and colleagues'finding [14].
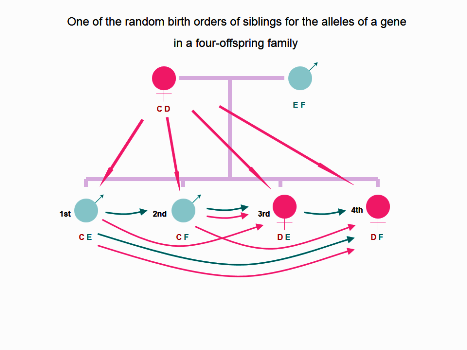
Figure 2. In one of 256 probabilities for the random birth order of siblings in an imagined fouroffspring family for the alleles of a gene, the long straight pink arrowhead indicates the prenatal MFCT,
and/or postnatal MICT via which, each offspring during fetal life and during lactation has chance to expose
to NIMAs encoded by noninherited maternal allele C, or D. The curved pink arrowhead indicates the
prenatal FFCT and postnatal FICT-mediated exposure to NIMAs carried by elder siblings. The curved blue
arrowhead indicates prenatal FFCT and postnatal FICT-mediated exposure to NIPAs carried by elder
siblings. The first born offspring cannot be exposed to the NIPA F despite of the exposure of NIMA D via
prenatal MFCT, postnatal MICT and FBBIC Cycle. The second born offspring may be exposed to NIPA E via
prenatal FFCT, postnatal FICT and FBBIC Cycle, in addition to the exposure of NIMA D via prenatal MFCT,
postnatal MICT and FBBIC Cycle. The third born offspring may be exposed to NIPA F via prenatal FFCT and
postnatal FICT from the second born brother and FBBIC Cycle. In addition, the third born girl may be
exposed to NIMA C via prenatal MFCT, postnatal MICT and FBBIC Cycle, as well as via prenatal FFCT and
postnatal FICT from the first and second born brothers. The forth born girl may be exposed to NIPA E via
prenatal FFCT and postnatal FICT from the first born brother and the third born sister and FBBIC Cycle.
The youngest girl may be exposed to NIMA C via prenatal FFCT and postnatal FICT from the first and the
second born brothers, and via prenatal MFCT, postnatal MICT and FBBIC Cycle.
Moreover, the inter-sibling tolerance between the first child and their younger
siblings in a family is influenced by birth order, while the inter-sibling tolerance among
those young siblings is mainly affected by the random birth orders that are dependent on
sibship size. For example, in the six-child family model (see Figure 1), there are 4096
total probabilities for the random birth orders of six siblings for the alleles of a gene. The
chance for the eldest offspring to be exposed to NIPA E or F for is zero, while it is 50%
(2048/4096) for the second child, 75% (3072/4096) for the third offspring, 87.5%
(3584/4096) for the fourth offspring, 93.75% (3840/4096) for the fifth one, and 96.88%
(3968/4096) for the youngest sibling, the sixth offspring.
However, it worth to point out three special orders: (i) all offspring in a family have
the same alleles of a gene for inherited paternal and maternal antigens. In other words,
there are no NIMAs and NIPAs among siblings, but NIMAs between maternal body and
offspring. Therefore, theoretically, prenatal FFCT and postnatal FICT and FBBIC Cycle
cannot offer a chance to each offspring to contact NIPAs, in contrast, each offspring may
contact NIMAs via prenatal MFCT and/or FFCT, and/or postnatal MICT, FICT and
FBBIC Cycle (see Figure 3), (ii) all offspring in a family have the same alleles for
inherited paternal antigens and different alleles for inherited maternal antigens (see
Figure 4), and (iii) all offspring have the same alleles for inherited maternal antigens and
different alleles for inherited paternal antigens (see Figure 5).
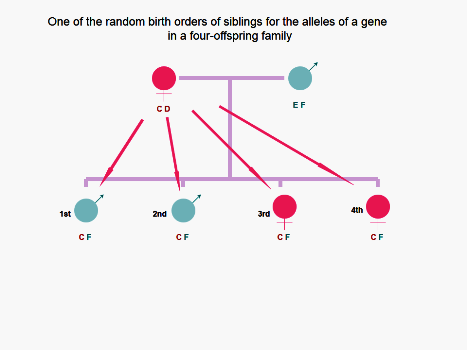
Figure 3. In the imagined 4-children family, one order (1st.CF, 2nd.CF, 3rd.CF, and 4th.CF) of the four
probabilities that the four offspring have the same inherited paternal and maternal antigens is illustrated.
Therefore, theoretically, prenatal FFCT and postnatal FICT and FBBIC Cycle cannot offer a chance to each of
offspring to contact NIPA E at the cellular level; in contrast, every offspring may contact NIMA D via
prenatal MFCT and/or FFCT, and/or postnatal MICT, FICT and FBBIC Cycle.
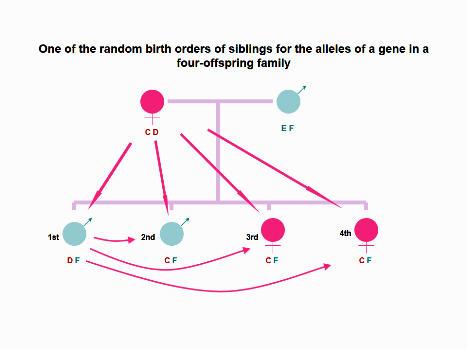
Figure 4. In the imagined 4-children family, four siblings have the same alleles for inherited paternal
antigens and the first born boy bears a different allele (D) of a gene for inherited maternal antigens
compared to his three younger siblings. Theoretically, the eldest son may contact NIMA C via only prenatal
MFCT and postnatal MICT, and every younger offspring may contact NIMA D via prenatal MFCT and FFCT,
and/or postnatal MICT, FICT and FBBIC Cycle. However, prenatal FFCT and postnatal FICT and FBBIC Cycle
cannot offer a chance to each of offspring to contact NIPA E at the cellular level.
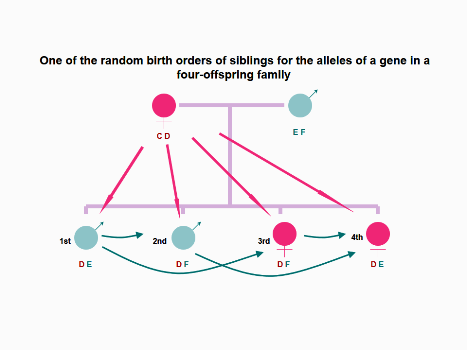
Figure 5. In the imagined 4-children family, four siblings have the same alleles for inherited maternal
antigens and the first born boy and the fourth born girl bear a different allele (E) of a gene for inherited
paternal antigens compared with the second and third born siblings. Theoretically, the second born boy and
the third born girl may contact NIPA E from their eldest brother via prenatal FFCT and postnatal FICT and
FBBIC Cycle. In contrast, the eldest brother was unable to contact NIPA F from the second and third born
younger siblings at the cellular level. The youngest girl may contact NIPA F from the second born brother
and the third born sister via prenatal FFCT and/or postnatal FICT and FBBIC Cycle. All four offspring can
contact NIMA C from maternal body via prenatal MFCT and postnatal MICT and FBBIC Cycle at the cellular
level.
Clearly, in the NIPAs-centric model, the exposure to NIPAs may occur only when
offspring of the different birth orders bear different alleles for inherited paternal antigens.
In contrast, the exposure to NIMAs can occur via prenatal MFCT and postnatal MICT
and FBBIC Cycle, even if all offspring share the same maternal allele (C or D).
When the hypothesized family has two-offspring, the random birth order of two
siblings for the alleles of a gene has total 16 probabilities, and the chance for the eldest
brother or sister to contact NIPA E or F via FFCT and FICT, or even FBBIC Cycle is
zero; in contrast, that for the second offspring to be exposed to either NIPA E or F is 50%
(8/16). Moreover, the chance for both the firstborn sibling and the second offspring to
match both NIMA and NIPA is 25% (4/16); and therefore, for the second-born sibling the
chance of tolerance to NIPA E or F is higher than that for the firstborn sibling.
In the family with three siblings, the random birth order of three siblings for the alleles
of a gene has a total of 64 probabilities. The chance for the firstborn offspring to contact
NIPA E or F via FFCT, FICT and FBBIC Cycle is zero; in contrast, the chance for the
second offspring to be exposed to either NIPA E or F is 50% (32/64); and that for the
third-born offspring, the youngest sibling, to contact NIPA E or F is as high as 75%
(48/64).
When sibship size increases to four, the random birth order of four siblings for the
alleles of a gene has a total of 256 probabilities. The chance for the firstborn brother or
sister to contact NIPA E or F is still zero; but the chance for the second offspring is 50%
(128/256), for the third is 75% (192/256), and for the fourth offspring, the youngest
sibling is 87.5% (224/256).
In a five-sibling family, the random birth order of five siblings for the alleles of a gene
has a total of 1024 probabilities. The chance to be exposed to NIPA E or F is zero for the
first offspring; 50% (512/1024) for the second one; 75% (768/1024) for the third
offspring; 87.5% (896/1024) for the fourth offspring, and 93.75% (960/1024) for the fifth
one, the youngest sibling.
In the six-sibling microchimeric model, the random birth order of six siblings for the
alleles of a gene has a total of 4096 probabilities. The chance to be exposed to NIPA E or
F for the eldest offspring is zero, 50% (2048/4096) for the second one, 75% (3072/4096)
for the third offspring, 87.5% (3584/4096) for the fourth offspring, 93.75% (3840/4096)
for the fifth one, and 96.88% (3968/4096) for the youngest sibling, the sixth offspring.
The possible reason for the Gratwohl and colleagues’ failure to show the birth order
effect [28] may be due to their simply division of 6062 cases into two groups: (1) donor
younger than recipient, and (2) donor older than recipient [28]. In fact, according to the
NIPAs-centric model, the chance to being exposed to NIPA E or F for a later-born sibling
increases as the sibling size increases.
Theoretically, the first living offspring in the hypothesized family should be the result
of the first pregnancy that the mother experienced, and under this strict definition, the
chance of being exposed to NIPA E or F for the first-born offspring is always zero.
However, in fact, many women may have experienced previously silent abortions or
unsuccessful pregnancies before having delivered their first living baby, which should be
given special attention in future laboratory and epidemiological studies.
V. Murine models in the experimental animal study on tolerogenic mechanism
It is worth to point out that theoretically, in multiple pregnancy animals, such as mice,
rats, and pigs, the chance of exposure to NIPA should be equal in every one of littermates
[29]. Therefore, multiple pregnancy animals may not be a suitable experimental model
for studying the birth order and sibship size effect in transplantation immunology.
In the experimental animal study, the murine model used by Andrassy and colleagues
was the product of H-2(b/b) males mating with H-2(b/d) females [13], which, however, is
suitable for studying the NIMAs mechanism but not for others because they are multiple
pregnancy mammals and theoretically, there should be a fetus-to-fetus cell transmission
or exchange during their normal gestation [29]. Therefore, the murine offspring of H-2(b/d) males mating with H-2(b/b) females may be suitable for studying NIPAstolerogenic mechanism, FFCT, and FICT.
The survival of grafts is a synthetic manifestation at the individual macroorganism
level, meaning the mechanisms involved in tolerance are complicated; and in fact, there
are still some critical questions to be answered. The NIMAs-centric model alone cannot
account for the birth order effect and the NIMAs paradox in transplantation immunology.
The novel theoretical model proposed in the manuscript shows that the variation of intersibling tolerance to NIPAs via FFCT and FICT is associated with birth order and sibship
size, and that the first-born child has no chance to contact NIPAs carried by his/her
younger siblings, and therefore, no tolerance to those NIPAs. In contrast, prenatal FFCT
and MFCT and postnatal FICT, MICT and FBBIC Cycle may offer later-born siblings the
chance to be exposed to NIPAs carried by their elder siblings or previously aborted
embryos/fetuses, and therefore, to induce tolerance to those NIPAs.
Abbreviations
FMCT: gestation-associated fetus-to-maternal cell transmission;
MFCT: gestation-associated maternal-to-fetal cell transmission;
FFCT: gestation-associated fetus-to-fetus cell transmission;
MICT: breastmilk and breastfeeding-mediated maternal-to-infant cell transmission;
FICT: breastmilk and breastfeeding-mediated transmission of sibling and allogenic fetal cells into infantile body;
FBBIC Cycle: fetus-to-infant his/her own fetal cell external transmission via breastfeeding;
FMMC: fetal-maternal microchimerism;
MFMC: maternal-to-fetal microchimerism;
FFMC: fetofetal microchimerism;
MIMC: maternal-infantile microchimerism;
FIMC: fetoinfantile microchimerism;
NIPAs: noninherited paternal antigens;
NIMAs: noninherited maternal antigens.
Reference
1. Billingham RE, Brent L, Medawar PB. Actively acquired tolerance of foreign cells. Nature. 1953; 172(4379): 603-6
2. Starzl TE. History of clinical transplantation. World J Surg. 2000; 24(7): 759-82
3. Mold JE, Michaelsson J, Burt TD, Muench MO, Beckerman KP, Busch MP, Lee TH, Nixon DF, McCune JM. Maternal alloantigens promote the development of tolerogenic fetal regulatory T cells in utero. Science. 2008; 322(5907): 1562-5
4. Burlingham WJ, Grailer AP, Heisey DM, Claas FH, Norman D, Mohanakumar T, Brennan DC, de Fijter H, van Gelder T, Pirsch JD, Sollinger HW, Bean MA. The effect of tolerance to noninherited maternal HLA antigens on the survival of renal transplants from sibling donors. N Engl J Med. 1998; 339(23): 1657-64
5. Tamaki S, Ichinohe T, Matsuo K, Hamajima N, Hirabayashi N, Dohy H, et al. Japan Society of Hematopoietic Cell Transplantation. Superior survival of blood and marrow stem cell recipients given maternal grafts over recipients given paternal grafts. Bone Marrow Transplant. 2001; 28(4): 375-80
6. Stern M, Ruggeri L, Mancusi A, Bernardo ME, de Angelis C, et al. Survival after T cell-depleted haploidentical stem cell transplantation is improved using the mother as donor. Blood. 2008; 112(7): 2990-5
7. Owen RD, Wood HR, Foord AG, Sturgeon P, Baldwin LG. Evidence for actively acquired tolerance to Rh antigens. Proc Natl Acad Sci USA. 1954; 40(6): 420-4
8. Claas FH, Gijbels Y, van der Velden-de Munck J, van Rood JJ. Induction of B cell unresponsiveness to noninherited maternal HLA antigens during fetal life. Science. 1988; 241(4874): 1815-7
9. Hadley GA, Phelan D, Duffy BF, Mohanakumar T. Lack of T-cell tolerance of noninherited maternal HLA antigens in normal humans. Hum Immunol. 1990; 28(4): 373-81
10. Roelen DL, van Bree FP, van Beelen E, van Rood JJ, Claas FH. No evidence of an influence of the noninherited maternal HLA antigens on the alloreactive T cell repertoire in healthy individuals. Transplantation. 1995; 59(12): 1728-33
11. Smits JM, Claas FH, van Houwelingen HC, Persijn GG. Do noninherited maternal antigens (NIMA) enhance renal graft survival? Transpl Int. 1998; 11(2): 82-8
12. Opelz G. The effect of tolerance to noninherited maternal HLA antigens on the survival of renal transplants from sibling donors. N Engl J Med. 1999; 340(17): 1369-70
13. Andrassy J, Kusaka S, Jankowska-Gan E, Torrealba JR, Haynes LD, Marthaler BR, Tam RC, Illigens BM, Anosova N, Benichou G, Burlingham WJ. Tolerance to noninherited maternal MHC antigens in mice. J Immunol. 2003; 171(10): 5554-61
14. Bucher C, Stern M, Buser A, Heim D, Paulussen M, Halter J, et al. Role of primacy of birth in HLA-identical sibling transplantation. Blood. 2007;110(1): 468-9
15. Dutta P, Molitor-Dart M, Bobadilla JL, Roenneburg DA, Yan Z, Torrealba JR, Burlingham WJ. Microchimerism is strongly correlated with tolerance to noninherited maternal antigens in mice. Blood. 2009; 114(17): 3578-87
16. Gratwohl A, Doehler B, Stern M, Bucher C, Passweg J, Opelz G. Birth order and outcome after HLA-identical sibling donor transplantation. Blood. 2009 114(27): 5569-70
17. Burlingham WJ, Benichou G. Bidirectional alloreactivity: A proposed microchimerism-based solution to the NIMA paradox. Chimerism. 2012; 3: 1-8
18. Ponsonby AL, Couper D, Dwyer T, Carmichael A. Cross section study of the relation between sibling number and asthma, hay fever, and eczema. Arch Dis Child. 1998; 79(4): 328-33
19. Karmaus W, Arshad H, Mattes J. Does the sibling effect have its origin in utero? Investigating birth order, cord blood immunoglobulin E concentration, and allergic sensitization at age 4 years. Am J Epidemiol. 2001; 154(10): 909-15
20. Karmaus W, Botezan C. Dose a higher number of siblings protect against the development of allergy and asthma? A review. J Epidemiol Community Health. 2002; 56(3): 209-17
21. McKeever TM, Lewis SA, Smith C, Collins J, Heatlie H, Frischer M, Hubbard R. Early exposure to infections and antibiotics and the incidence of allergic disease: a birth cohort study with the West Midlands General Practice Research Database. J Allergy Clin Immunol. 2002; 109(1): 43-50
22. Bernsen RM, de Jongste JC, van der Wonuden JC. Birth order and sibship size as independent risk factors for asthma, allergy, and eczema. Pediatr Allergy Immunol. 2003; 14(6): 464-9
23. Beelen DW, Ottinger HD, Elmaagacli A, Scheulen B, Basu O, Kremens B, et al. Transplantation of filgrastim-mobilized peripheral blood stem cells from HLA-identical sibling or alternative family donors in patients with hematologic malignancies: a prospective comparison on clinical outcome, immune reconstitution, and hematopoietic chimerism. Blood. 1997; 90(12): 4725-35
24. Weisdorf DJ, Anasetti C, Antin JH, Keman NA, Kollman C, Snyder D, et al. Allogenic bone marrow transplantation for chronic myelogenous leukemia: comparative analysis of unrelated versus matched sibling donor transplantation. Blood. 2002; 99(6): 1971-7
25. Ottinger HD, Ferencik S, Beelen DW, Lindemann M, Peceny R, Elmaagacli AH, Husing J, Grosse-Wilde H. Hematopoietic stem cell transplantation: contrasting the outcome of transplantations from HLA-identical siblings, partially HLA-mismatched related donors, and HLA-matched unrelated donors. Blood. 2003; 102(3):1131-7 (PMID: 12689945)
26. Perez-Garcia A, De la Camara R, Torres A, Gonzalez M, Jimenez A, Gallardo D. Minor histocompatibility antigen HA-8 mismatch and clinical outcome after HLA-identical sibling donor allogeneic stem cell transplantation. Haematologica. 2005; 90(12):1723-4 (PMID: 16330460)
27. van Rood JJ, Scaradavou A, Stevens CE. Indirect evidence that maternal microchimerism in cord blood mediates a graft-versus-leukemia effect in cord blood transplantation. Proc Natl Acad Sci USA. 2012; 109(7): 2509-14
28. Gratwohl A, Doehler B, Stern M, Bucher C, Passweg J, Opelz G. Birth order and outcome after HLA-identical sibling donor transplantation. Blood. 2009; 114(27): 5569-70
29. Yin S-d. Six models of the pregnancy-associated eukaryotic cell transmission among fetus, mother, and infant. The Journal of Theoretical Fimpology. 2013, 1(2): e-20120609-1-2-3. Available from:
http://www.fimpology.com30. Li XC, Wells AD, Strom TB, Turka LA. The role of T cell apoptosis in transplantation tolerance. Curr Opin Immunol 2000; 1(5):522-7
31. Starzl TE. Chimerism and tolerance in transplantation. Proc Natl Acad Sci USA. 2004; 101(Suppl 2): 14607-14
32. Mathew JM, Leventhal JR, Miller J. Microchimerism in promoting graft acceptance in clinical transplantation. Curr Opin Organ Transplant. 2011; 16(4): 345-52
33. Lambert NC, Pang JM, Yan Z, Erickson TD, Stevens AM, Furst DE, Nelson JL. Male microchimerism in women with systemic sclerosis and healthy women who have never given birth to a son. Ann Rheum Dis. 2005; 64(6): 845-8
34. Yan Z, Lambert NC, Guthrie KA, Porter AJ, Loubiere LS, Madeleine MM, Stevens AM, Hermes HM, Nelson JL. Male microchimerism in women without sons: quantitative assessment and correlation with pregnancy history. Am J Med. 2005; 118(8): 899-906
35. McConico A, Butters K, Lien K, Knudsen B, Wu X, Platt JL, Ogle BM. In utero cell transfer between porcine littermates. Reprod Fertil Dev. 2011; 23(2): 297-302
36. Karniychuk UU, Van Breedam W, Van Roy N, Rogel-Gaillard C, Nauwynck HJ. Demonstration of microchimerism in pregnant sows and effects of congenital PRRSV infection. Vet Res. 2012; 43(1): 19
37. Molitor ML, Haynes LD, Jankowska-Gan E, Mulder A, Burlingham WJ. HLA class I noninherited maternal antigens in cord blood and breast milk. Hum Immunol. 2004; 65(3): 231-9
38. Aoyama K, Matsuoka KI, Teshima T. Breast milk and transplantation tolerance. Chimerism. 2010; 1(1): 19-20
Copyright © 2013- by Cory H. E. R. & C. Inc. All Rights Reserved.